DUAL-KLAS-NIR
Version:
P700, Plastocyanin, Ferredoxin & Chlorophyll Fluorescence Measuring System




What Does the DUAL-KLAS-NIR Do?
The photosynthetic electron transport chain passes electrons from water through photosystem II, plastoquinone, the cytochrome b6f complex, plastocyanin, P700 (the photosystem I reaction center), and ferredoxin to NADP⁺. Understanding how electrons flow through this chain and where bottlenecks occur is key to understanding photosynthetic regulation, stress responses, and the coordination between the two photosystems. The DUAL-KLAS-NIR is the only instrument that can monitor the redox state of P700, plastocyanin, and ferredoxin simultaneously and in real time. By adding chlorophyll fluorescence data from the integrated PAM fluorometer, the system provides a comprehensive picture of the entire electron transport chain in a single measurement - from PSII to the acceptor side of PSI.
The DUAL-PAM-100 employs one pair of wavelengths in the near-infrared (NIR) to measure absorbance changes related to redox changes of the photosystem I reaction center. Using four wavelength pairs in the NIR (780/820, 820/870, 870/965 and 840/965 nm), the DUAL-KLAS-NIR is capable of unequivocally discriminating redox changes of plastocyanin, P700, and ferredoxin. The software of the DUAL-KLAS-NIR shares many features with that of the DUAL-PAM-100, making the transition straightforward for existing users. Using automated measuring routines, even complex measuring protocols can be easily performed.
How Does the Deconvolution Work?
The redox changes of P700, plastocyanin, and ferredoxin all produce overlapping absorbance changes in the near-infrared. By applying an innovative analytical approach, the DUAL-KLAS-NIR acquires the in vivo spectral characteristics of pure PC, P700, and Fd. This spectral information allows the system to monitor online the redox changes of all three components and to determine PC/P700 and Fd/P700 ratios, thereby providing direct estimates of the relative pool sizes of these electron carriers.
Integrated PAM Chlorophyll Fluorometer
The DUAL-KLAS-NIR includes a full PAM chlorophyll fluorometer with both green and blue measuring light. Green light penetrates deeper into the leaf than blue light - thus, green-excited fluorescence includes information from deeper leaf layers and provides the best match with the NIR absorbance measurements, which always probe the entire leaf. The blue measuring light gives more specific information on the upper or lower leaf side and produces a fluorescence signal that is one order of magnitude larger than the green-excited signal.
Flexible Time Resolution
The system measures in 1, 2, or 6 channel mode with time resolutions of 35 µs, 150 µs, and 1 ms respectively. The pulse modulation technique developed for the DUAL-PAM-100, in which channels are measured as blocks of 50 µs, has been extended for the DUAL-KLAS-NIR to accommodate 6-channel measurements. An extremely wide range of measuring light frequencies (1 Hz to 400 kHz) supports both continuous F0 assessment and high-resolution recording of fast kinetic transients such as the polyphasic fluorescence rise or flash relaxation kinetics.
Versatile Light Sources
Integrated red (635 nm), blue (460 nm), and far-red actinic LEDs provide continuous actinic illumination, multiple turnover pulses, and single turnover flashes. The red LED array serves as the main actinic light source, while the blue LED in the detector unit allows continuous illumination with up to 300 µmol m⁻² s⁻¹ from the detector side of the sample. Both sides of the leaf can be illuminated for homogeneous light distribution or, by removing the LED array cable of the DKN-E emitter unit, only one side. All light sources switch with 2.5 µs time resolution under software control.
Single-Channel High-Resolution Mode
When single-channel measurements are made, a dedicated script allows the different signals to be measured consecutively at high time resolution combining the speed of single-channel mode with the information content of multi-channel measurements.
Automated Measurement Protocols
The software supports automated slow kinetics recordings, pre-programmed triggered runs, online and offline averaging, and operation via automated measuring routines through script file programming. This enables reproducible, multi-step experiments without manual intervention.


The DUAL-KLAS-NIR is unique in providing online deconvoluted signals of the in vivo redox states of P700, PC and Fd in real time. This opens up a whole new research field with applications that may not have come to our minds yet. In particular, for the first time simultaneous kinetic information on the complex interplay between reactions at the PS I donor and acceptor sides can be obtained, including information on cyclic electron transport around PS I.
Some examples of applications, which have been described in Klughammer and Schreiber, Photosynth Res 128 (2016) 195-214, Schreiber and Klughammer, Plant Cell Physiol 57 (2016) 1454-1467 and Schreiber Photosynth Res 134 (2017) 343-360, are given here.

The effective quantum yield of photosystem I. In all older measurements the P700 signal contained always contributions of Fd and variable amounts of PC, depending on the method used to reduce this contribution. Panel C of the figure illustrates the light intensity dependence of the effective PSI quantum yield Y(I), the effective quantum yield of PSII Y(II) and the effective quantum yield of PSII corrected for PSI fluorescence Y(II)corr of Brassica napus.


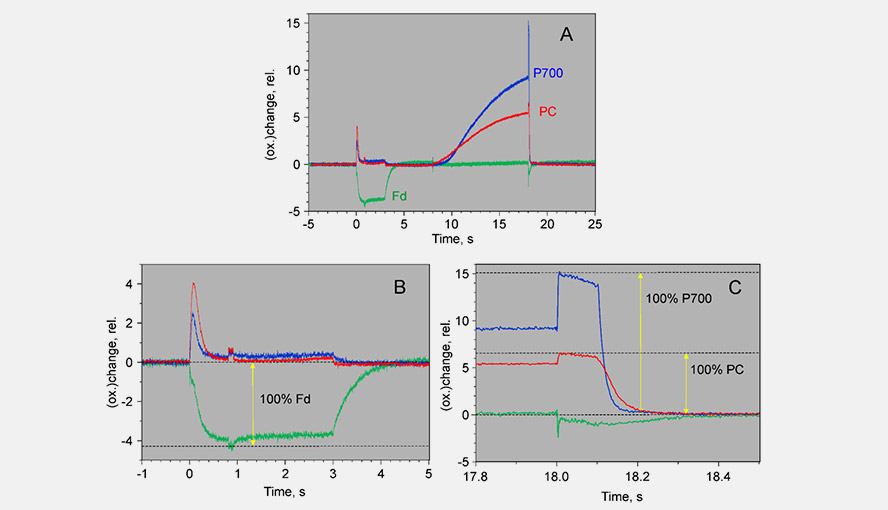
The effective antenna size of photosystem I. On a dark-to-light transition, first oxidized PC starts to accumulate and only with some delay oxidized P700. The initial slope of the clean PC signal can now be used as a measure for the effective antenna size of PS I. The second figure zooms in on the initial absorbance changes and shows the large difference in the initial slope of PC (red) and P700 (blue). For a dark-acclimated leaf, with inactive photosystem I acceptor side, the initial slope of Fd-reduction can also be used for this purpose. The figure illustrates the delay in the oxidation kinetics of P700 relative to those of PC for a barley leaf, which has a relatively high PC/P700 ratio.

The DUAL-KLAS-NIR has a window to visualize the simultaneous changes in the P700 and PC redox states relative to each other. This allows an estimation of the apparent equilibrium constant between PC and P700. The figure gives an example of a P700 versus PC redox plot.


During a saturation pulse given to a well dark-adapted leaf, the Fd-pool becomes reduced and in a subsequent period of darkness slowly re-oxidizes again. When the electron flow on the acceptor side of PSI has become activated the re-oxidation kinetics become much faster. Following activation of the acceptor side of PSI, the dark-inactivation can be followed by monitoring the rate at which Fd becomes re-oxidized following a probe pulse. These kinetics can be fit with an exponential fit routine. Panel A gives examples of the Fd reoxidation kinetics fit with an exponential fit routine and Panel B shows the dark-inactivation kinetics of the acceptor side of PSI for a Hedera helix leaf.


The maximal NIR transmittance changes of PC, P700 and Fd are proportional to the leaf/sample content of these compounds and the ratios of the extinction coefficients of PC, P700 and Fd are constant. This makes it possible to probe the PC/P700 and Fd/P700 ratios and thereby the relative PC and Fd pool sizes in different species or under different conditions (e.g. sun/shade, stressed/non-stressed) on a routine basis with the DUAL-KLAS-NIR. It is observed that high PC/P700 ratios correlate with high ETR values. The figure illustrates that sun and shade leaves of, in this case, Hedera helix have considerably different PC and Fd contents relative to P700.










Scientific Publications using Walz Devices
Source: Google Scholar.
Keywords: (Walz OR Waltz) Effeltrich.
Date: June 22, 2026.
Ʃ = 19642
Source: Google Scholar.
Keywords: (Walz OR Waltz) Effeltrich.
Date: June 22, 2026.
Ʃ = 19642
Selected Publications
Effect of the insecticide clothianidin on the photosynthetic electron transport chain in pea.
Photochemistry and Photobiology 101: 580-591
Plastocyanin affects photosynthesis and high light acclimation by modulating redox states of electron transport chain in Chlamydomonas reinhardtii
Communications Biology 8: 476
Role of plastocyanin in response to photosynthetic electron transport in Solanum nigrum under Cd stress
Plant Stress 18: 101073
Metabolite-level regulation of enzymatic activity controls awakening of cyanobacteria from metabolic dormancy
Current Biology 35: 77-86
The impact of redox mediators on the electrogenic and physiological properties of Synechocystis sp. PCC 6803 in a biophotovoltaic system
ChemSusChem 18: e202402543
Unravelling the physiological and anatomical basis of divergent adaptations in cultivated and wild tomatoes
Journal of Experimental Botany 76: 6548-6566
Molecular dynamics of photosynthetic electron flow in a biophotovoltaic system
Environmental Science and Ecotechnology 23: 100519
Towards solar chemicals: Understanding and redesigning photosynthesis
Thesis University Turku
Moderate temperature reduction changes the high-light acclimation strategy of lettuce plants
Physiologia Plantarum 177: e70298
Sub-optimal temperature leads to tighter coupling between photosynthetic electron transport and CO2 assimilation under fluctuating light in maize
bioRxiv-2
The regulation of PSI cyclic electron transport by both plastoquinone and ferredoxin redox states. correlation with the rate of proton motive force utilization
Frontiers in Plant Science 16: 1626163
PSII photoinhibition as a protective strategy: maintaining an oxidative state of PSI by suppressing PSII activity under environmental stress
Physiologia Plantarum 177: e70392
The protective role of chloroplast NADH dehydrogenase-like complex (NDH) against PSI photoinhibition under chilling stress
New Phytologist 248: 2262-2279
The role of the LysR-type transcriptio factor PacR in regulating nitrogen metabolism in Anabaena sp. PCC7120
Physiologia Plantarum 177: e70248
Redox state of photosynthetic ferredoxin under heat and light stress.
Journal of Applied Spectroscopy 91: 342-348
Glycogen synthesis prevents metabolic imbalance and disruption of photosynthetic electron transport from photosystem II during transition to photomixotrophy in Synechocystis sp. PCC 6803.
New Phytologist 243: 162-179
Dynamics and interplay of the photosynthetic regulatory processes depend on the amplitudes of oscillating light.
Plant, Cell & Environment 47: 2240-2257
A mathematical model of photoinhibition: exploring the impact of quenching processes.
in silico Plants 6: diae001
Flavonols do not affect aphid load in green or senescing birch leaves but coincide with a decrease in photosystem II functionality.
Biology Open 13: bio060325
Strong heterologous electron sink outcompetes alternative electron transport pathways in photosynthesis.
The Plant Journal 119: 2500-2513
Manganese deficiency alters photosynthetic electron transport in Marchantia polymorpha.
Plant Physiology and Biochemistry 215: 109042
A mathematical model to simulate the dynamics of photosynthetic light reactions under harmonically oscillating light.
Plant Physiology and Biochemistry 217: 109138
Proton gradient regulation 5 is required to avoid photosynthetic oscillations during light transitions.
Journal of Experimental Botany 75: 947-961
Flv3A facilitates O2 photoreduction and affects H2 photoproduction independently of Flv1A in diazotrophic Anabaena filaments.
New Phytologist 237: 126-139
Evaluating the oxidation rate of reduced ferredoxin in Arabidopsis thaliana independent of photosynthetic linear electron flow: plausible activity of ferredoxin-dependent cyclic electron flow around photosystem I.
International Journal of Molecular Sciences 24: 12145
Plants cope with fluctuating light be frequency-dependent nonphotochemical quenching and cyclic electron transport.
New Phytologist 239: 1869-1886
Eukaryote-specific assembly factor DEAP2 mediates an early step of photosystem II assembly in Arabidopsis.
Plant Physiology 193: 1970-1986
The photosystem I supercomplex from a primordial green alga Ostreococcus tauri harbors three light-harvesting complex trimers.
eLife12: e84488
Enhanced function of non-photoinhibited photosystem II complexes upon PS II photoinhibition.
BBA-Bioenergetics 1864: 148978
Higher reduced state of Fe/S-signals, with the suppressed oxidation of P700, causes PS I inactivation in Arabidopsis thaliana.
Antioxidants 12: 21
High cyclic electron transfer via the PGR5 pathway in the absence of photosynthetic control.
Plant Physiology 192: 370-386
Scrutinizing the impact of alternating electromagnetic fields on molecular features of the model plant Arabidopsis thaliana.
International Journal of Environmental Research and Public Health 19: 5144
Determining Photosynthetic Control, a probe for the balance between electron transport and Calvin-Benson cycle activity, with the DUAL-KLAS-NIR.
Photosynthesis Research 153: 191-204
CP12 fine-tunes the Calvin-Benson cycle and carbohydrate metabolism in cyanobacteria.
Frontiers in Plant Science 13: 1028794
Insight on the regulation of photosynthesis in pea leaves exposed to oscillating light.
Journal of Experimental Botany 73: 6380-6393
Plants acclimate to photosystem I photoinhibition by readjusting the photosynthetic machinery.
Plant, Cell & Environment 10: 2954-2971
Synechocystis sp. PCC 6803 requires the bidirectional hydrogenase to metabolize glucose and arginine under oxic conditions.
Frontiers in Microbiology 13: 896190
Evidence for electron transfer from the bidirectional hydrogenase to the photosynthetic complex I (NDH-1) in the Cyanobacterium Synechocystis sp. PCC 6803.
Microorganisms 10: 1617
Flv3A facilitates O2 photoreduction and affects H2 photoproduction independently of Flv1A in diazotrophic Anabaena filaments.
bioRxiv
Pyruvate:ferredoxin oxidoreductase and low abundant ferredoxins support aerobic photomixotrophic growth in cyanobacteria.
bioRxiv
Photosynthetic linear electron flow drives CO2 assimilation in maize leaves.
International Journal of Molecular Sciences 22: 4894
PGRL2 triggers degradation of PGR5 in the absence of PGRL1.
Nature Communications 12: 3941
Singlet oxygen, flavonols and photoinhibition in green and senescing silver birch leaves.
Trees 35: 1267–1282
Dynamic thylakoid stacking and state transitions work synergistically to avoid acceptor-side limitation of photosystem I.
Nature Plants 7: 87-98
Developmental acclimation of the thylakoid proteome to light intensity in Arabidopsis.
The Plant Journal 105: 223–244
Differences in ionic, enzymatic, and photosynthetic features characterize distinct salt tolerance in eucalyptus species.
Plants 10: 1401
In-vivo quantification of electron transport makes up about 35% in a cyanobacterium.
Biochimica et Biophysica Acta 1862: 148353
redundancy and crosstalk between flavodiiron proteins and NDH-1 in Synechocystis sp. PCC 6803.
BioRxiv
Redox regulation by peroxiredoxins is linked to their thioredoxin-dependent oxidase function.
Photosynthesis Research 145: 31-41
Near-infrared in vivo measurements of photosystem I and its luminal electron donors with a recently developed spectrophotometer.
Photosynthesis Research 144: 63-72
Identification of the electron donor tot flavodiiron proteins in Synechocystis sp. PCC 6803 by in vivo spectroscopy.
Biochimica et Biophysica Acta 1861: 148256
Functional redundancy between flavodiiron proteins and NDH-1 in Synechoscystis sp. PCC 6803.
The Plant Journal 103: 1460-1476
Intrinsic fluctuations in transpiration induce photorespiration to oxidize P700 in photosystem I.
Plants 9: 1761
A commonly used photosynthetic inhibitor fails to block electron flow to Photosystem I in intact systems.
Frontiers in Plant Science 11: 382
Interference between arsenic-induced toxicity and hypoxia.
Plant, Cell & Environment 42: 574-590
Oxidation of P700 induces alternative electron flow in photosystem I in wheat leaves.
Plants 8: 152
Consequences of photosystem-I damage and repair on photosynthesis and carbon use in Arabidopsis thaliana.
Plant Journal 97: 1061-1072
Multilevel regulation of non-photochemical quenching and state transitions by chloroplast NADPH-dependent thioredoxin reductase.
Physiologia Plantarum 166: 211-225
Near-infrared in vitro measurements of photosystem I cofactors and electron-transfer partners with a recently developed spectrophotometer.
Photosynthesis Research 142: 307-319
Regulation of cyclic electron flow by chloroplast NADPH-dependent thioredoxin system.
Plant Direct 2: e00093
Changing frequency of fluctuating light reveals the molecular mechanism for P700 oxidation in plant leaves.
Plant Direct 2: e00073
PROTON GRADIENT REGULATION 5 supports linear electron flow to oxidize photosystem I.
Physiologia Plantarum 164: 337–348
The chloroplast 2-cysteine peroxiredoxin functions as thioredoxin oxidase in redox regulation of chloroplast metabolism.
eLife 7: e38194
Redox changes of ferredoxin, P700, and plastocyanin measured simultaneously in intact leaves.
Photosynthesis Research 134: 343–360
Deconvolution of ferredoxin, plastocyanin, and P700 transmittance changes in intact leaves with a new type of kinetic LED array spectrophotometer.
Photosynthesis Research 128: 195–214
Analysis of photosystem I donor and acceptor sides with a new type of online-deconvoluting kinetic LED-array spectrophotometer.
Plant and Cell Physiology 57: 1454-1467
DUAL-KLAS-NIR
Windows computer (Windows 7/8/10/11) with KLAS-100 software
Base plate, 40 cm x 30 cm
73.5 cm, diameter 1.5 cm
2.8 kg
Accessories
Black-anodized aluminum body with central 10 x 10 mm glass cuvette; for attachment of Measuring Heads and Miniature Magnetic Stirrer PHYTO-MS; additional ports for attachment of two additional measuring heads
750 g
Based on a device manufactures by h+p (type Variomag-Mini); featuring adapter to be mounted in the bottom port of the Optical Unit ED-101US/MD; powered and controlled by the Power-and-Control-Unit
3.7 mm Ø diffusing sphere coupled to integrated PAR sensor via 2 mm diameter fiber; compact amplifier unit and special holder for mounting on Optical Unit ED-101US/MD; to be connected tot the Power-and-Control Unit
Magnetic stirrer driven by a rotating magnetic field; the PHYTO-MS is connected to Power-and-Control Unit PHYTO-II-C; a special adapter plug allows the insertion in the bottom port of the Optical Unit ED-101US/MP
Power-and-Control Unit US-T/DR
Peltier Heat-Transfer Head US-T/DS
0.29 kg (including cable)
AC Adapter
100 V - 240 V AC 1.5 A 50-60 Hz
130 mm x 56 mm x 30 mm (L x W x H)
Pneumatically separated upper and lower cuvette halves. Controlled by a regulator unit with sockets for cable connections to the Control Unit 3000 C of the GFS-3000 and a trigger input line.
General Features and Graphical User Interface
The KLAS-100 software used for DUAL-KLAS-NIR measurements is written in the tradition of the DUAL-PAM-100 software. It allows the user to make the sample quickly ready for online signal deconvolution and the measurements the user has in mind via a fixed sequence of steps. A script is provided to determine the maximum P700, PC and Fd signal amplitudes (comparable to the Pm determination of the DUAL-PAM-100). Four NIR wavelength pairs are measured and the P700, PC and Fd contributions to these NIR signals are determined using so-called differential model plots or DMPs. Differential Model Plots for Hedera helix are included in the software for reference. In practice, separate sets of DMPs have to be determined for individual plant species. Via a fixed sequence of steps, making use of the scripts included in the software for the determination of each DMP (P700, PC and Fd), the user can make his or her own set of DMPs for the photosynthetic organism that is studied.
The software has a window for saturation pulse analysis and one for light curves, just like in the DUAL-PAM-100 software, but then including an analysis of the PC and Fd redox states. Like for the DUAL-PAM-100, the software automatically calculates classical fluorescence ratio parameters as well as more recently suggested parameters. In a new window it is possible to plot the P700 and PC redox states against each other, which gives an idea about the redox-equilibrium between the two. Trigger files can be created and Script files written that in combination allow the reproducible execution of complex experimental protocols. Whereas some background knowledge is required for programming Trigger files and Scripts, even very sophisticated protocols can be reliably executed by non-experts. The software gives the user the flexibility to create almost any experimental protocol that can be imagined for the analysis of electron transport related processes.










Differential Model Spectra (DMPs) for the deconvolution of the NIR-signals
The absorption spectra of P700, plastocyanin (PC) and ferredoxin (Fd) are broad and show few features in the near infrared (NIR). A further complication is that the extinction coefficient of P700 is much higher than that of PC, which is higher again than the extinction coefficient of Fd. By choosing specific wavelength pairs it is only possible to get NIR-signals enriched in Fd (780-820 nm), or P700 (820-870 nm) or PC (870-950 nm). To get from there to a clean deconvolution, the software makes use of the differential model plot (DMP) approach, which does not require knowledge of difference spectra and the wavelength dependence of differential extinction coefficients. Christof Klughammer first developed this method for the KLAS-100, a kinetic LED array spectrophotometer for the 510-570 nm wavelength range.
We know the behavior of PC, P700 and Fd under standard conditions quite well and we can, on the basis of that knowledge, design experiments in which, for a short period of time, only redox changes of one of the three components (PC, P700 or Fd) occur. By normalization of the values determined for the 4 difference signals a “spectral fingerprint” for each component is obtained under the given experimental conditions. It should be noted here as well that the FA and FB iron-sulfur centers located on the acceptor side of photosystem I may also contribute to the Fd-signal.



Usage
Highlights
- The only instrument that separates P700, plastocyanin, and ferredoxin signals in real time
- Four NIR wavelength pairs for unambiguous deconvolution of overlapping absorbance changes
- Integrated PAM chlorophyll fluorometer with green and blue measuring light
- Time resolution down to 35 µs in single-channel mode
Downloads
- Software: KLAS-100_update_v3_0_16
- Manuals & Documentation: DUAL-KLAS-NIR Manual